 手機版
手機版 化工儀器網(wǎng)手機版
化工儀器網(wǎng)手機版
 化工儀器網(wǎng)小程序
化工儀器網(wǎng)小程序
 官方微信
官方微信 公眾號:chem17
公眾號:chem17
 掃碼關注視頻號
掃碼關注視頻號
")











趨化因子及其受體的功能

免疫細胞的定向遷移是機體免疫應答發(fā)生和完成的必須條件,。趨化因子是一類控制細胞定向遷移的細胞因子,。其功能行使由趨化因子受體介導。趨化因子與其受體的相互作用控制著各種免疫細胞在循環(huán)系統(tǒng)和組織器官間定向遷移, 使之到達感染,、創(chuàng)傷和異常增殖部位, 執(zhí)行清除感染源,、促進創(chuàng)傷愈合和消滅異常增殖細胞、維持組織細胞平衡的功能,。趨化因子及其受體的表達和功能異常,,將導致免疫細胞不能在正確的位置行使正確的功能。因此趨化因子系統(tǒng)在免疫系統(tǒng)功能行使的各個環(huán)節(jié)中處于關鍵地位,,并由此在病原體的清除,、炎癥反應、病原體感染,、細胞及器官的發(fā)育,、創(chuàng)傷的修復、腫瘤的形成及其轉移,、移植免疫排斥等方面都起著重要的作用,。以趨化因子及其受體分子為控制靶點,通過激活或拮抗趨化因子受體的信號傳導來調控趨化因子系統(tǒng)的功能,,可望用于控制和治療相關疾病,。
- 趨化因子與趨化因子受體
趨化因子是一類能趨化細胞定向移動的小分子分泌蛋白,由70至100個氨基酸組成,。至今已發(fā)現(xiàn)了40多種人的趨化因子,,屬細胞因子中的zui大家族。當成纖維細胞,、內(nèi)皮細胞,、表皮細胞等組織細胞和免疫細胞在受到刺激物如生長因子、干擾素、病毒產(chǎn)物及細菌產(chǎn)物的誘導時可分泌出不同的趨化因子,。在趨化因子的分子中都有4個保守的半胱氨酸(C),。根據(jù)靠近分子氨基端(N端)的前兩個C間是否插入其它氨基酸, 將它們分為4個亞類:CXC類(插入1個氨基酸殘基),亦稱為α類趨化因子,,如IL-8(Interleukin-8);CC 類(不插入其它氨基酸殘基),又稱為β類趨化因子, 如MCP-1(Monocyte chemoattractant protein-1);CX3C類(插入3個其它氨基酸),,如Fractalkine;C類(N端僅1個C),如Lymphotactin ,。目前所發(fā)現(xiàn)的趨化因子主要屬于α類和β類,。在分子結構上,皆通過二硫鍵折疊成以自由的N-端,、三個反向折疊的β-片層和α螺旋的羧基端(C-端)為特征的二級結構,。
趨化因子受體是一類介導趨化因子行使功能的GTP-蛋白偶連的跨膜受體(GPCR),通常表達于免疫細胞,、內(nèi)皮細胞等細胞膜上,。分子由約330個氨基酸組成。7個跨膜區(qū)將分子分成細胞外自由的N-端,、3個細胞外環(huán),、3個細胞內(nèi)環(huán)和C-端幾個部分。胞內(nèi)第二環(huán)是與異三聚體G-蛋白偶連的部位,,有特征的天門冬氨酸-精氨酸-酪氨酸盒(DRY box)氨基酸序列。與趨化因子受體偶連的異三聚體G-蛋白的α亞基為Gi/o,,對百日咳毒素敏感,。按趨化因子的分類, 將同CC 類趨化因子結合的受體稱為CC類受體(CCR), 同CXC類趨化因子結合的受體稱為CXC 類受體(CXCR),,同樣有C和CX3C受體(CR,、CX3CR)。
趨化因子N-端氨基酸殘基的缺失突變體不能與其受體結合, 證實了趨化因子的N-端序列在與受體結合及引起信號轉導中的關鍵作用,。C端能夠極大地增強N端肽段的信號轉導功能,。1st -beta片層結構能夠和血管內(nèi)皮細胞上的氨基葡聚糖GAG(Glycosaminoglycan)結合, 可使其附著并富集于血管內(nèi)皮細胞上。趨化因子受體胞外區(qū)的N端及一個以上的胞外環(huán)參與其配體的結合, 而且N端的序列在很大程度上決定了受體對趨化因子的特異選擇性,。趨化因子受體的C端(胞內(nèi)部分)大都為富含絲氨酸(Serine)和蘇氨酸(Threonine)殘基的片段,,其磷酸化可能參與受體活化后的信號傳導和內(nèi)化。
在配體-受體結合實驗中, 可以看到趨化因子與受體間結合的冗余現(xiàn)象, 即一個趨化因子可與數(shù)個趨化因子受體結合,,如RANTES(Regulated on activation normal T cell expressed and secreted)與CCR1,、CCR3、CCR5的結合,;一個趨化因子受體可與數(shù)個趨化因子結合,,如CCR3 與eotaxins , MCP-2 、3 、4 ,、RANTES的結合,。因此, 在體外趨化實驗中表現(xiàn)為。一種趨化因子可以趨化表達不同趨化因子受體的免疫細胞做定向遷移,一種免疫細胞可以為多種趨化因子所趨化,。正是這種趨化因子及其受體相互作用的冗余才使趨化因子系統(tǒng)在體內(nèi)的精細調控成為可能,。在體內(nèi),各種趨化因子通過它們在組織中不同時相的表達和分布差異, 以及趨化因子受體在不同免疫細胞類群上不同時相的表達和分布差異, 調控著不同免疫細胞的定向遷移和相互作用, 并由此決定了趨化因子與其受體作用的特異性,。如IL-8主要作用于中性粒細胞的定向遷移,;CXCR3、CCR5的表達局限在T輔助1型細胞(TH1)上,,由INF-gamma可誘導產(chǎn)生的MIG(Monokine induced by INF-γ),、IP-10(INF-γinducible protein of Mr 10 000)及I-TAC(INF-inducible T cell αchemoattractant)主要分布在TH1型炎癥組織中;CCR3,,CCR4,,CCR8 則局限在T輔助2型細胞(TH2)上;TARC(Thymus- and activation-related chemokine)表達于皮膚血管內(nèi)皮上,,向皮膚組織聚集的T細胞表達其受體CCR4,;小腸固有層血管內(nèi)皮表達TECK(Thymus-expressed chemokine),在腸炎組織中有表達其受體CCR9的T細胞浸潤,。
- 趨化因子對免疫細胞的趨化作用
趨化因子的基本功能就是對表達有相應趨化因子受體的細胞的定向趨化作用,。目前認為, 免疫細胞克服血管內(nèi)皮細胞屏障,在體液和組織間穿行包括4個步驟, 即細胞隨體液流動,、細胞被穩(wěn)固黏附到血管內(nèi)皮上,、細胞穿過內(nèi)皮細胞間隙、細胞遷移到特定組織中,。在此過程中, 趨化因子控制著滲出細胞的選擇性以及被選擇細胞的穩(wěn)固黏附,。體外實驗已證實趨化因子能使在血流速度下的淋巴細胞黏附在固相支持物上, 不同的趨化因子特異地引導表達相應趨化因子受體的淋巴細胞的附著。免疫細胞由于所表達的選擇素與血管內(nèi)皮細胞上的選擇素受體的相互作用,,與血管內(nèi)皮有瞬間的非選擇性的可逆性黏附,,因此免疫細胞在血流中沿血管壁做滾動前行。局部組織中血管內(nèi)皮細胞所分泌的趨化因子通過內(nèi)皮細胞上的GAG被富集在血管內(nèi)皮表面,。表達相應趨化因子受體的免疫細胞在滾動前行中由于與血管內(nèi)皮上趨化因子作用而促使免疫細胞整合素的上調,,整合素與內(nèi)皮細胞上的黏附分子的相互作用導致免疫細胞不可逆地黏附到血管內(nèi)皮表面。穩(wěn)固黏附的免疫細胞在其分泌的特殊酶的作用下,,穿過內(nèi)皮細胞間隙和基底膜,,并在趨化因子濃度梯度的引導下,移行至特定組織中,。
- 趨化因子與天然免疫
研究表明, 趨化因子具有直接抑菌作用,。趨化因子MIG,、IP-10及I-TAC在體外抑菌實
驗中具有防御素樣的直接抑菌作用。經(jīng)IFN-γ刺激PBMC所產(chǎn)生的MIG,、IP-10及I-TAC是非刺激細胞的28 ~ 35倍, 在瓊脂擴散實驗中能夠抑制大腸桿菌和李斯特菌的生長,。而且, 與防御素相同,該抑菌作用受高濃度NaCl的抑制,。另外,,人β-防御素是趨化因子受體CCR6
的配體,能趨化表達CCR6的未成熟樹突狀細胞(Dendritic cell ,DC)和記憶性T細胞的定向遷移, 由此可以推測,,天然免疫因子β-防御素通過與CCR6的相互作用, 招募DC和T細胞到達微生物感染部位, 是啟動獲得性免疫應答的途徑之一,。
- 趨化因子與獲得性免疫
4 .1抗原提呈與T、B 細胞活化
DC是目前發(fā)現(xiàn)的功能zui強的抗原提呈細胞(Antigen presenting cell ,APC),。DC從外周組織遷移到周圍淋巴組織伴隨著其抗原提呈過程亦即DC的成熟過程,。趨化因子通過作用于具有不同趨化因子受體表達譜的DC,控制和調節(jié)DC完成其遷移過程,。未成熟DC表達多種趨化因子受體, 在感染和創(chuàng)傷組織中不同的趨化因子的招募下進入損傷組織,,在進行抗原攝取和處理的過程中, 由不表達CCR7 的未成熟DC 轉變?yōu)楦弑磉_CCR7的成熟DC(mDC),使之能在表達于輸入淋巴管高內(nèi)皮靜脈(HEV)和次級淋巴組織中的副皮質區(qū)(T-區(qū))的趨化因子SLC ELC (Secondary lymphoid tissue chemokine Epstein-Barr virus-induced receptor ligand chemokine)的引導下,,從周圍組織進入二級淋巴組織中,。同樣, 幼稚T細胞亦高表達CCR7。與mDC一樣,受HEV和周圍淋巴組織中SLC/ELC的引導進入淋巴結的副皮質區(qū)(T-區(qū)域), 并在此接受APC提呈的抗原信息而被激活成效應T細胞,。效應T細胞離開淋巴結并經(jīng)血液循環(huán)游走到外來抗原部位啟始特異性免疫反應,。幼稚(Naive)B細胞亦高表達CCR7和CXCR5,受SLC/ELC和表達于周圍淋巴組織淺皮質區(qū)的BCA-1(B cell-activating chemokine-1)的引導,,通過HEV進入淋巴結并定位于淺皮質區(qū)(B細胞區(qū)域)而被活化,。
4 .2 T 細胞的極化
T 細胞極化成TH1或TH2代表著T 細胞應答的重要特征。環(huán)境中的細胞因子類型決定了T細胞極化的方向,。IL-12、INF-α促使幼稚T細胞極化成TH1細胞,;IL-4則促使稚T細胞極化成TH2細胞,;而TGF-β能夠促使幼稚T細胞發(fā)展成TH0細胞,后者具有被極化成TH1細胞或TH2細胞的潛能,。極化的T細胞表達不同的趨化因子受體, 從而使不同的T細胞被趨化到不同的*位,,發(fā)揮不同的生物學功能。TH1細胞特征表達CCR5,、CXCR3,,受趨化因子如MIP-1α(Macrophage inflammatory protein-1α)、RANTES,、MIG 和IP-10 的驅使到達損傷組織, 并能表達上述趨化因子以募集更多的淋巴細胞,,引起TH1型炎癥反應;TH2細胞特征表達CCR3、CCR4,、CCR8,,可以受趨化因子如eotaxin-1、2,、3, MCP-2,、3、4的驅使而到達損傷組織,,引發(fā)TH2型炎癥反應,。趨化因子能通過誘導TH0細胞定向分化來調節(jié)前炎性細胞因子與抗炎細胞因子的平衡。MIP-1α與活化的T細胞孵育可導致向TH1的極化,,增加IFN-γ的表達水平,,下調IL-4的水平;而MCP-1與活化的T細胞孵育后則導致向TH2的極化,,增加IL-4的水平,,下調IFN-γ水平。zui近還發(fā)現(xiàn), IP-10,、MIG,、I-TAC既是TH1細胞特征表達的趨化因子受體CXCR3的激動劑,同時又是TH2表達的趨化因子受體CCR3的天然拮抗劑,,這一事實表明通過CXCR3招募TH1細胞的趨化因子做為CCR3的拮抗劑,,又能阻斷CCR3的趨化因子對TH2細胞的招募。因此,,趨化因子能促使不同極化T細胞的募集,。
4 .3病原體的清除
免疫活性細胞到達病原體感染部位是免疫系統(tǒng)進行抑制和清除病原體的先決條件。機體對結核分支桿菌的感染所產(chǎn)生的細胞免疫反應需要巨噬細胞和T細胞聚集到感染部位,。CCR2基因敲除小鼠在結核分支桿菌感染的早期就會死亡, 其感染肺部組織中細菌數(shù)量是正常感染小鼠的100倍,,在感染早期和晚期皆可觀察到巨噬細胞和DC及T細胞積聚到感染肺部的數(shù)量大大減少,表明CCR2及其相應趨化因子在啟始和控制結核分支桿菌感染中的重要作用,。
- 趨化因子的其他功能
趨化因子遠非僅僅是zui初認為的機體的免疫調節(jié)因子,,還明確地具有除調節(jié)免疫細胞遷移以外的其它功能, 如一些趨化因子是細胞生長調節(jié)因子,一些是血管生成調節(jié)因子,,一些則參與胚胎期免疫系統(tǒng),、循環(huán)系統(tǒng)和中樞神經(jīng)系統(tǒng)的形成和發(fā)育,還有一些趨化因子受體是一些病原體入侵宿主細胞的受體,。這些發(fā)現(xiàn)將趨化因子及其受體的重要作用擴展到參與免疫系統(tǒng)功能以外的生物學功能之中,。
5 .1調節(jié)血細胞發(fā)育
趨化因子系統(tǒng)以多種方式來調節(jié)血細胞的發(fā)育。*,,可以調節(jié)造血干細胞和血細胞前體的增殖,。體內(nèi)外實驗證實,,有幾種趨化因子如MIP-1α、IL-8 ,能夠抑制正常血細胞前體的增殖,,并有協(xié)同作用,。由于這種抑制作用能夠保護造血細胞免受抗腫瘤藥物導致的對細胞周期中造血前體細胞的毒性作用,MIP-1α的類似物BB10010做為治療乳腺癌化療中血細胞減少的藥物進入Ⅱ期臨床試驗,。SDF-1能促進人外周血CD34+前體細胞的增殖,,增強人骨髓、脊柱血及小鼠脊柱中造血干細胞的生存和抗凋亡能力, 增強體外長期培養(yǎng)小鼠造血干細胞的植入能力,。第二,,可以調節(jié)造血干細胞和血細胞前體的遷移和歸巢。在SDF-1/CXCR4基因敲除小鼠上觀察到骨髓中造血前體細胞而非胎肝中造血前體細胞減少, 表明SDF-1在胚胎發(fā)育中造血干細胞和前體細胞從胎肝向骨髓的遷移中起決定性作用,。第三,,調節(jié)淋巴細胞發(fā)育中淋巴細胞的遷移和歸巢。許多趨化因子組成性地表達于胸腺組織,、周圍淋巴結和其它淋巴組織中,,選擇性地招募表達有相應趨化因子受體的幼稚淋巴細胞,使得胸腺及周圍淋巴組織能夠發(fā)育正常。這些趨化因子被稱為歸巢趨化因子(Homing chemokines),。如在胸腺組織中表達有趨化因子TECK,、ELC、SLC,、MIP-3α等, 在二級淋巴組織中表達有ELC ,、SLC、DCCK-1(Dendritic cell chemokine-1),、BCA-1等,,招募幼稚T細胞和B 細胞分別進入各自的發(fā)育成熟部位。
5 .2參與胚胎期器官的發(fā)育
SDF-1/CXCR4基因敲除小鼠死于子宮內(nèi),伴有嚴重的心室隔膜缺損,小腦組織結構發(fā)育異常,胃腸道的血管發(fā)育異常,顯示它們在器官發(fā)育中的重要作用,。
5 .3趨化因子與血管生成
在CXC類趨化因子中,就其CXC 前至N-端是否存在ELR(Glu-Leu-Arg)序列將之分為2個亞類, 即ELR+-CXC 和ELR-CXC,。ELR+-CXC 類趨化因子,如IL-8、MGSA(Melanoma growth-stimulatory activity), 能趨化培養(yǎng)的內(nèi)皮細胞定向移動, 并能增強家兔角膜新生血管的數(shù)量, 因此具有促血管生成作用;而ELR-CXC類趨化因子,如IP-10,、MIG,、PF4(Plaet factor 4),則具有抑制血管生成作用, 它們在體外趨化試驗中能夠拮抗ELR+-CXC 類趨化因子對內(nèi)皮細胞的趨化作用,,并抑制ELR+-CXC類趨化因子的促家兔角膜血管生成作用。促血管生成功能在促進創(chuàng)傷的愈合和腫瘤的生長中有重要作用,。
5 .4趨化因子受體與病原微生物感染
5 .4 .1瘧原蟲受體
瘧原蟲入侵紅細胞是通過紅細胞上的Duffy 抗原受體(Duffy antigen receptor for chemokines,,DARC)介導。DARC是紅細胞上能夠結合多種趨化因子的趨化因子受體,,是發(fā)現(xiàn)的*個可以和不同類趨化因子結合(如可以和CC類的MCP-1,、RANTES及CXC類的IL-8,、MGSA結合)的趨化因子受體,但不能引發(fā)信號轉導,。在功能上可能是血液中的趨化因子池,,以吸收并調節(jié)血液循環(huán)中過量的趨化因子。
5 .4 .2 HIV的輔助受體
HIV感染宿主細胞除需要細胞上的CD4為受體外, 還需要趨化因子受體做為其輔助受體才能實現(xiàn),。HIV首先與宿主細胞上的CD4 結合,,進一步與相應的趨化因子受體CCR5或CXCR4結合,zui終導致HIV核酸穿膜進入宿主細胞,。主要的輔助受體是CCR5 和CXCR4,。由于分離時病毒所利用的輔助受體的差別,將HIV分為嗜巨噬細胞株(M-株)和嗜T細胞株(T-株), 前者主要侵染細胞表面表達有CCR5的巨噬細胞,,后者主要侵染細胞表面表達有CXCR4的T淋巴細胞,。通常在初始感染者上分離到的HIV都是M-株。由于HIV的高變異性,,在持續(xù)感染一段時間后,,從感染者身上還可以分離到能夠利用其它的趨化因子受體的
HIV,如CCR2,、CCR3,、CXCR2等。參與輔助受體功能的結構主要是趨化因子受體的N-端,、胞外區(qū)的一些部位,,它們與其趨化因子結合區(qū)域有部分交叉重疊。
5 .5 趨化因子對組織細胞及腫瘤細胞的趨化作用
趨化因子除了對免疫細胞有定向趨化作用外,,對其它能夠表達相應趨化因子受體的組織細胞都具有趨化作用,。如前面提到的ELR-CXC類趨化因子對內(nèi)皮細胞的趨化作用。SDF-1對胚胎期神經(jīng)細胞的趨化能力使之在中樞神經(jīng)系統(tǒng)神經(jīng)網(wǎng)絡的發(fā)育中起重要作用,。長期以來一直有報道,,在腫瘤組織中有趨化因子和趨化因子受體的表達,包括腫瘤細胞和浸潤細胞,。但趨化因子對腫瘤的發(fā)生和發(fā)展的正向或負向作用是多方面的和不確切的,。一些趨化因子通過激活免疫活性細胞毒性細胞或抑制腫瘤相關血管生成來抑制腫瘤的生長和發(fā)展。而另一些趨化因子則通過促腫瘤細胞增生,、移行,,促進蛋白水解酶的分泌和誘導血管生成來促進腫瘤的發(fā)展。在不同的研究中, 不同的趨化因子有不同的作用,,或相同的趨化因子也有不同的作用, 表明在腫瘤的形成和發(fā)展中趨化因子網(wǎng)絡的參與是復雜的,。腫瘤的轉移是一種非隨機的、有組織器官選擇性的高度組織化的過程,,與淋巴細胞的定向遷移有相似之處,。已有實驗證實腫瘤細胞的轉移受趨化因子的嚴格調控,。與正常乳腺細胞和乳腺組織相比,在乳腺癌培養(yǎng)細胞,、乳腺癌組織和乳腺癌轉移瘤上有高表達CCR7和CXCR4,。CCR7的配體SLC/ELC表達于HEV和周圍淋巴組織中,CXCR4的配體SDF-1主要表達于淋巴結、肺,、肝及骨髓, 而在小腸,、腎、大腦和骨骼肌中痕跡量表達,。乳腺癌轉移特征正是向周圍淋巴結,、骨髓、肺及肝臟轉移, 極少至腎臟,。在體外實驗中,,相應趨化因子對乳腺癌細胞有趨化作用和誘導浸潤作用;體內(nèi)實驗表明, 抗CXCR4的單抗能夠阻止乳腺癌細胞轉移至淋巴結和肺組織。同樣,,在黑色素瘤的組織特異性轉移與腫瘤細胞上表達的趨化因子受體CXCR4,、CCR7 和CCR10 有關。因此趨化因子及其受體在決定腫瘤轉移目地組織中有重要作用,。
- 結語
趨化因子及其受體的上述生物學功能的研究和揭示使之成為當今zui有前途的治療炎癥性疾病的靶點,。趨化因子及其受體相互作用的多渠道方式可能意味著細胞在移行的每一步驟都依賴于單個的趨化因子或趨化因子受體, 阻斷這一受體可導致該移行細胞移行方向的改變, 所以在許多趨化因子受體拮抗劑的實驗研究中可以看到, 往往單個趨化因子的阻斷就能明顯地抑制炎癥反應。不同的淋巴細胞在趨化因子的作用下選擇性地移行到特定的部位,,提示阻斷趨化因子受體策略較目前所使用的免疫抑制劑更具選擇性和安全性,。目前已在制備各種趨化因子受體拮抗劑, 有單克降抗體、趨化因子變異體和小分子有機化合物,。在未來的幾年中很有可能就會從中篩選到治療免疫相關疾病的有效藥物,。
相關通路指標:
Chemokine Ligands:
α Chemokine (CXC Motif): CXCL1 (IL8RA), CXCL10 (INP10), CXCL11 (I-TAC/IP-9), CXCL12 (SDF1), CXCL13, CXCL14, CXCL16, CXCL2 (IL8RB), CXCL3, CXCL5 (ENA-78/LIX), CXCL6 (GCP2), CXCL9 (MIG).
β Chemokine (CC Motif): CCL1 (I309), CCL11 (Eotaxin), CCL13 (MCP-4), CCL14, CCL15 (MIP-1D), CCL16 (HCC4), CCL17 (TARC), CCL18 (PARC), CCL19, CCL2 (MCP-1), CCL20 (MIP-3A), CCL21 (MIP-2), CCL22 (MDC), CCL23 (MPIF-1), CCL24 (Eotaxin 2), CCL25 (TECK), CCL26 (Eotaxin 3), CCL27, CCL28, CCL3 (MIP-1A), CCL4 (MIP-1B), CCL5 (RANTES), CCL7 (MCP-3), CCL8 (MCP-2).
γ Chemokine (C Motif): XCL1, XCL2.
δ Chemokine (CX3C Motif): CX3CL1.
Chemokine Receptors:
α Chemokine (CXC Motif): CXCR1, CXCR2, CXCR3, CXCR4, CXCR5, CXCR6.
β Chemokine (CC Motif): CCR1, CCR10, CCR2, CCR3, CCR4, CCR5, CCR6, CCR7, CCR8, CCR9.
γ Chemokine (C Motif): XCR1.
δ Chemokine (CX3C Motif): CX3CR1.
Atypical: ACKR2, ACKR4, CCRL2, ACKR3, DARC.
Cytokines Involved in Chemotaxis: C5, CKLF, CMTM1 (CKLFSF1), CMTM2 (CKLFSF2), CMTM3, CMTM4, IL16, IL1B, IL4, IL8, PF4V1, PPBP, TNF.
Cytokine Receptors Involved in Chemotaxis: CR1 (GPR77), CMKLR1, FPR1, GPR17.
Other Genes Involved in Chemotaxis: HIF1A, SLIT2, TLR2, TLR4, TYMP.
通路研究工具:
癌癥炎癥因子和免疫衰減系列 | PCR Array of Cancer Inflammation & Immunity Crosstalk | HU-CIIC-084 |
趨化因子受體系列 | PCR Array of Chemokines & Receptors | HU-CHKR-084 |
克羅恩病系列 | PCR Array of Crohn's Disease | HU-CD-084 |
樹突狀抗原呈遞細胞系列 | PCR Array of Dendritic & Antigen Presenting Cell | HU-DAPC-084 |
G蛋白偶聯(lián)受體系列 | PCR Array of G Protein Coupled Receptors | HU-GPCR-084 |
炎性細胞因子及受體系列 | PCR Array of Inflammatory Cytokines & Receptors | HU-ICR-084 |
炎癥反應和自身免疫系列 | PCR Array of Inflammatory Response & Autoimmunity | HU-IRA-084 |
PCR Array:
 Q-PCR array是信號通路基礎生物學研究和臨床疾病研究的重要工具。這種功能分類芯片運用成熟的SYBR Green熒光定量PCR技術,,專注于一個信號通路中基因的表達水平檢測,。與傳統(tǒng)的高密度表達譜芯片相比,Q-PCR array具有針對性強,、靈敏度高,、準確可靠等優(yōu)點。 芯片上的基因包括了與研究對象有確定關系的基因,,或至少是與研究對象的關系有待考證的基因,。如果研究對象是某一生物學通路的基因,則使用針對該類基因的功能分類基因芯片比使用高密度表搭配芯片更加有效便利,。
Q-PCR array是信號通路基礎生物學研究和臨床疾病研究的重要工具。這種功能分類芯片運用成熟的SYBR Green熒光定量PCR技術,,專注于一個信號通路中基因的表達水平檢測,。與傳統(tǒng)的高密度表達譜芯片相比,Q-PCR array具有針對性強,、靈敏度高,、準確可靠等優(yōu)點。 芯片上的基因包括了與研究對象有確定關系的基因,,或至少是與研究對象的關系有待考證的基因,。如果研究對象是某一生物學通路的基因,則使用針對該類基因的功能分類基因芯片比使用高密度表搭配芯片更加有效便利,。
BioTNT QPCR芯片特點:
l 靈敏度高,,樣本使用量低;
l 線性范圍廣,可同時檢測表達水平差異大的基因,;
l 每個基因的檢測引物優(yōu)化后保證高擴增率和產(chǎn)物單一;
l 重復性高,,Ct值的平均差異只有0.25個循環(huán),;
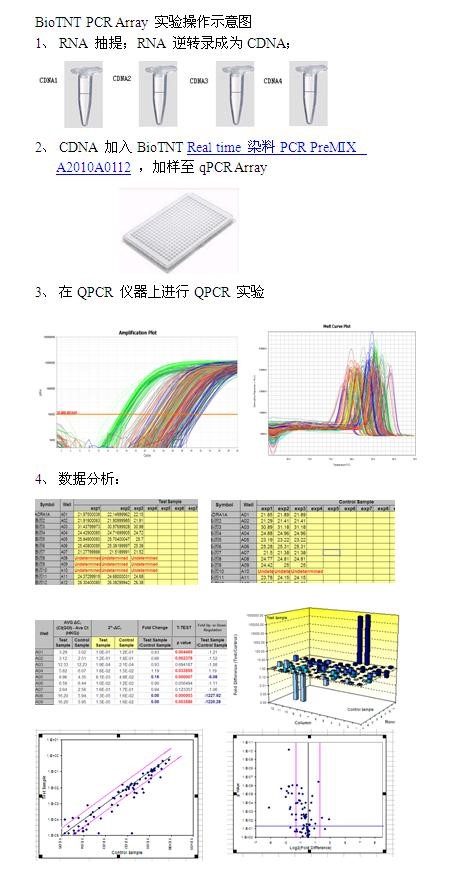
BioTNT QPCR Array芯片操作流程:
相關產(chǎn)品




免責聲明
- 凡本網(wǎng)注明“來源:化工儀器網(wǎng)”的所有作品,均為浙江興旺寶明通網(wǎng)絡有限公司-化工儀器網(wǎng)合法擁有版權或有權使用的作品,,未經(jīng)本網(wǎng)授權不得轉載,、摘編或利用其它方式使用上述作品。已經(jīng)本網(wǎng)授權使用作品的,,應在授權范圍內(nèi)使用,,并注明“來源:化工儀器網(wǎng)”。違反上述聲明者,,本網(wǎng)將追究其相關法律責任,。
- 本網(wǎng)轉載并注明自其他來源(非化工儀器網(wǎng))的作品,目的在于傳遞更多信息,,并不代表本網(wǎng)贊同其觀點和對其真實性負責,,不承擔此類作品侵權行為的直接責任及連帶責任。其他媒體,、網(wǎng)站或個人從本網(wǎng)轉載時,,必須保留本網(wǎng)注明的作品第一來源,并自負版權等法律責任,。
- 如涉及作品內(nèi)容,、版權等問題,請在作品發(fā)表之日起一周內(nèi)與本網(wǎng)聯(lián)系,,否則視為放棄相關權利,。


 采購中心
采購中心