 手機(jī)版
手機(jī)版 化工儀器網(wǎng)手機(jī)版
化工儀器網(wǎng)手機(jī)版
 化工儀器網(wǎng)小程序
化工儀器網(wǎng)小程序
 官方微信
官方微信 公眾號(hào):chem17
公眾號(hào):chem17
 掃碼關(guān)注視頻號(hào)
掃碼關(guān)注視頻號(hào)
械、包裝設(shè)備與材料中國(guó)展 | 實(shí)驗(yàn)室儀器與裝備展區(qū)")
驗(yàn)儀器設(shè)備有限公司")










CD4+ Th細(xì)胞激活后分化為功能不同的Th1和Th2效應(yīng)細(xì)胞,Th1細(xì)胞分泌IL2,、IFNγ、TNFβ等介導(dǎo)細(xì)胞免疫應(yīng)答,、遲發(fā)型超敏反應(yīng)和器官特異自身免疫性疾病在宿主抗胞內(nèi)病原感染中起重要作用,。Th2 細(xì)胞產(chǎn)生IL4、IL5,、IL6、IL9,、IL10,、IL13等細(xì)胞因子介導(dǎo)體液免疫應(yīng)答、過(guò)敏性和感染性疾病在拮抗胞外病原體(如細(xì)菌,、寄生蟲(chóng)) ,、B 細(xì)胞增殖分化以及哮喘病等方面具有重要作用。近年來(lái),,人們對(duì)初始CD4 +T細(xì)胞分化為Th1或Th2細(xì)胞的機(jī)制進(jìn)行了廣泛而深入的研究,,已取得一些共同的認(rèn)識(shí):雙信號(hào)刺激活化CD4+T細(xì)胞,活化信號(hào)傳向胞內(nèi),、啟動(dòng)多條信號(hào)轉(zhuǎn)導(dǎo)通路,激活轉(zhuǎn)錄因子,、繼而轉(zhuǎn)錄因子結(jié)合到目的基因的啟動(dòng)子區(qū)、促使原癌基因,、細(xì)胞因子基因及其受體基因表達(dá),。細(xì)胞因子可經(jīng)自分泌和旁分泌作用促使T細(xì)胞克隆擴(kuò)增,,之后分化為功能各異的Th1和Th2 效應(yīng)細(xì)胞,部分分化為記憶細(xì)胞,。在Th細(xì)胞分化過(guò)程中,細(xì)胞因子被認(rèn)為是zui主要的因素,。
- 、TCR 和協(xié)同刺激信號(hào)
1.1TCR- CD3
TCR與MHCⅡ:抗原肽復(fù)合分子的結(jié)合為T細(xì)胞激活所需的*信號(hào),。TCR2CD3 激活后,CD3 的免疫受體酪氨酸活化基序( Immune receptor tyrosinebased activation motifs,,ITAM) 可轉(zhuǎn)導(dǎo)胞外刺激信號(hào)。研究發(fā)現(xiàn),在無(wú)IL4和IL12的T細(xì)胞分化早期,TCR 活化信號(hào)影響Th1PTh2 的分化決定:MEKPERK激活,、Ca2 +流增加驅(qū)使T細(xì)胞向Th1 分化,;PKC激活或鈣調(diào)磷酸酶部分阻斷則驅(qū)使T細(xì)胞向Th2分化。另外,,弱TCR 活化信號(hào)能激活Ca2 +流信號(hào)誘導(dǎo)IL4合成,,促使T細(xì)胞向Th2分化;強(qiáng)TCR活化信號(hào)可激活MAPK通路誘導(dǎo)IFN2γ合成,,促使T細(xì)胞向Th1分化,。TCR 觸發(fā)時(shí)間的長(zhǎng)短也影響Th細(xì)胞的分化;IL12存在時(shí),,短暫的TCR觸發(fā)啟動(dòng)Th1分化,長(zhǎng)時(shí)間的TCR觸發(fā)則啟動(dòng)Th2 分化,。
1.2 CD4
CD4 為T細(xì)胞的輔助受體,在T細(xì)胞活化早期能與MHCⅡ類(lèi)分子結(jié)合,,并通過(guò)蛋白酪氨酸激酶LcK轉(zhuǎn)導(dǎo)信號(hào),,LcK缺失的CD4+T細(xì)胞可出現(xiàn)Th2分化障礙,Tec家族激酶Rik 和Itk (Lck 下游分子)在Th2分化過(guò)程中亦起重要作用。
1.3 CD28及其同系物
CD28 與活化APC 上的B7.1 (CD80) 或B7.2(CD86)的結(jié)合為T細(xì)胞活化提供第二個(gè)信號(hào),。IL4,、IL5 以及IL10的產(chǎn)生需要CD28 協(xié)同刺激,CD28能增加胞內(nèi)轉(zhuǎn)錄因子NF-AT2 (nuclear factor of activated T cells 2)的累積,缺失時(shí)CD4 + T細(xì)胞僅分泌IFNγ。B7分子敲除的APC實(shí)驗(yàn)研究亦證實(shí)IL4 的產(chǎn)生和Th2分化高度依賴(lài)于B7 ,而有實(shí)驗(yàn)發(fā)現(xiàn)CD28能導(dǎo)致膜脂質(zhì)微小區(qū)域的重組和持續(xù)的酪氨酸磷酸化,但CD28介導(dǎo)Th2分化的分子機(jī)制目前尚不清楚,。zui近,Skapenko等發(fā)現(xiàn)記憶T細(xì)胞CD28 能啟動(dòng)IL4基因轉(zhuǎn)錄,并激活PI3激酶,、JNK-SAPK以及p38 MAP通路,CD28-B7誘導(dǎo)Th2分化要依賴(lài)于IL4的表達(dá)、p38 以及ERK-MAP通路的激活,。因此,CD8可能可以放大誘導(dǎo)Th2分化的TCR 活化信號(hào),。
CTLA4是CD28的同系物,表達(dá)于活化T細(xì)胞,與B7結(jié)合的親和力是CD28的10倍,,缺乏CTLA4可出現(xiàn)多克隆T細(xì)胞激活增殖紊亂,。CTLA4敲除T細(xì)胞能分泌高濃度IL4和IL5,提示CTLA4 在Th2分化誘導(dǎo)中起下調(diào)CD28的作用,。
誘導(dǎo)性協(xié)作刺激因子( inducible costimulator ,ICOS) 是近來(lái)發(fā)現(xiàn)的CD28同系物,亦表達(dá)于活化T細(xì)胞,。與CD28 和CTLA4 不同的是, ICOS 不與B7結(jié)合,而與B7RP1 (B7 的同系物之一) 結(jié)合。ICOS缺失小鼠T細(xì)胞分泌的IL4明顯降低,而IFNγ未見(jiàn)異常。因此,,ICOS 能潛在地調(diào)節(jié)T 細(xì)胞的分化發(fā)展,。
眾多來(lái)源的IL4 能通過(guò)STAT6 誘導(dǎo)Th2分化,但zui初驅(qū)使分化的IL4 源自何處,目前尚不清楚。IL4 基因定位于11號(hào)染色體,此部位還有IL5,、IL13 基因,。在IL4 和IL13 基因位點(diǎn)處發(fā)現(xiàn)了與Th2分化相關(guān)的幾個(gè)DNA 酶I高敏感位點(diǎn),提示:在Th2分化時(shí)伴隨有這些Th2型細(xì)胞因子基因位點(diǎn)的染色質(zhì)重組。已證實(shí)核小體上組蛋白的高乙?;cIL4,、IL5、IL13等基因相關(guān),且這種Th2特異的高乙?;蕾?lài)于STAT6 和GATA3 ,并伴隨IL4,、IL13基因的轉(zhuǎn)錄表達(dá)。目前在IL13 基因位點(diǎn)上游1. 6kbp處, 發(fā)現(xiàn)了一段保守的GATA3 反應(yīng)元件(GATA3 response element ,CGRE) 序列,它能與GATA3,、組蛋白乙?;D(zhuǎn)移酶復(fù)合物以及RNA 聚合酶Ⅱ結(jié)合,并具有增強(qiáng)上述Th2 型細(xì)胞因子基因啟動(dòng)子的作用。通過(guò)對(duì)IL4 基因啟動(dòng)子的研究,亦發(fā)現(xiàn)多個(gè)與IL4 基因表達(dá)有關(guān)的轉(zhuǎn)錄因子,如NFAT2AP1,、GATA3,、cmaf以及JunB 等。另外, IL13 能與IL4 受體α鏈結(jié)合,調(diào)節(jié)Th2 分化,; IL26 能上調(diào)細(xì)胞因子信號(hào)抑制分子1 ( suppressor of cytokine signaling1 ,SOCS1) 表達(dá)而抑制Th1 分化,同時(shí)促進(jìn)活化CD4+T 細(xì)胞分泌IL4 誘導(dǎo)Th2 分化,。
3.1 NF2AT
NF2AT家族有5個(gè)成員:NF2AT1 (NF2ATp) 、NF2AT2 (NF2ATc) ,、NF2AT3,、NF2AT4、NF2AT5 ,均有高度保守的DNA 和鈣調(diào)磷酸酶結(jié)合位點(diǎn),。前4者位于胞漿中,NF2AT5 則位于胞核內(nèi),。鈣調(diào)磷酸酶磷酸化介導(dǎo)NF2AT 的核內(nèi)轉(zhuǎn)運(yùn),而RasPRaf2MEK2ERK等蛋白激酶的激活使得NF2AT 與AP21 結(jié)合,形成NFAT2AP1復(fù)合物與DNA 結(jié)合。許多表達(dá)于活化T 細(xì)胞的基因啟動(dòng)子或增強(qiáng)子處均存在NF2AT 結(jié)合位點(diǎn),如IL2,、IL4,、IL25、IFNγ 等,。敲除NF2AT2 的小鼠,IL4 等Th2 型細(xì)胞因子表達(dá)受抑制,呈現(xiàn)Th2 分化缺陷,敲除NF2AT1 或同時(shí)敲除NF2AT1 和NF2AT4 小鼠卻出現(xiàn)截然相反的結(jié)果,。這說(shuō)明NF2AT1,、NF2AT2 能與IL4 啟動(dòng)子的NFAT2AP1 位點(diǎn)結(jié)合,功能各異地調(diào)節(jié)IL24 轉(zhuǎn)錄,。Porter 等報(bào)道, ERK1、JNK3,、p38α參與了NF2AT的磷酸化調(diào)節(jié),并能抑制NF2AT2的核轉(zhuǎn)運(yùn),。
3.2 GATA23
GATA23 屬于GATA 家族,表達(dá)于初始CD4+ T 細(xì)胞和朝Th2分化的Th0細(xì)胞,是Th2 分化調(diào)節(jié)的重要轉(zhuǎn)錄因子。過(guò)度表達(dá)GATA23 的轉(zhuǎn)基因小鼠T 細(xì)胞能表達(dá)IL4、IL5,、IL6,、IL10 和IL13 等Th2 型細(xì)胞因子mRNA,通過(guò)反義技術(shù)抑制GATA3 表達(dá),這些Th2 型細(xì)胞因子mRNA 的表達(dá)亦受抑制,。然而,IL4 基因近側(cè)啟動(dòng)子區(qū)并不存在GATA23 結(jié)合位點(diǎn),因此有人認(rèn)為GATA23 可能是一個(gè)Th2 特異的增強(qiáng)子,或是作為一個(gè)局部控制區(qū)域(如在IL4,、IL5、IL13 區(qū)域) 存在,。
在STAT6 敲除的T 細(xì)胞中, GATA23 亦能誘導(dǎo)Th2 型細(xì)胞因子表達(dá),導(dǎo)致Th2 分化,并在IL4 基因定位處發(fā)現(xiàn)Th2 特異的DNA 酶I 高敏感位點(diǎn);同時(shí),GATA3 能與NF2AT 結(jié)合而誘導(dǎo)IL5 啟動(dòng)子的活性,。近來(lái)研究發(fā)現(xiàn)一種稱(chēng)為FOG ( Friend of GATA) 的結(jié)構(gòu)分子,該分子表達(dá)于初始Th 細(xì)胞,起抑制GATA23 的作用。
3.3 c2maf
c2maf 是基本區(qū)域P亮氨酸拉鏈(basic region/leucine zipper) 轉(zhuǎn)錄因子,特異表達(dá)于Th2 細(xì)胞,是zui早被克隆的Th2 特異性轉(zhuǎn)錄因子,。以往認(rèn)為,c2maf激活后不影響其它Th2 型細(xì)胞因子的表達(dá),只是通過(guò)選擇性地與IL24 近側(cè)啟動(dòng)子結(jié)合,誘導(dǎo)IL24 表達(dá)促使Th2 分化,。zui近,哈佛醫(yī)學(xué)院的研究人員發(fā)現(xiàn),c2maf 亦可通過(guò)IL22Rα(CD25) 介導(dǎo)但不依賴(lài)IL4 的途徑促使Th2 分化。c2maf 滅活或敲除時(shí),Th2 分化和相應(yīng)細(xì)胞因子分泌照常進(jìn)行,推測(cè)可能與IL13的補(bǔ)償作用有關(guān),。
3.4 T-bet
T-bet (亦稱(chēng)T-box 21) 能激活IFNγ表達(dá),是新近發(fā)現(xiàn)的Th1 特異性轉(zhuǎn)錄因子,。T-bet 能誘導(dǎo)IFN-γ等位基因染色質(zhì)重組和轉(zhuǎn)錄激活IFN-γ基因,亦能誘導(dǎo)IL12Rβ2亞單位表達(dá),促使Th1 分化。若將T2bet導(dǎo)入已分化的CD4 Th2 細(xì)胞,出現(xiàn)IFN2γ分泌增強(qiáng),IL4,、IL5 等Th2 型細(xì)胞因子產(chǎn)生受阻,細(xì)胞朝Th1分化,。研究發(fā)現(xiàn),T2bet 誘導(dǎo)的Th1 分化不依賴(lài)于IL12-PSTAT4 或IL18 , 但受IL4-PSTAT6 的抑制。然而,STAT 與T2bet 之間的關(guān)系目前尚不清楚,。
3.5 CPEBP 和JunB
與c2maf 類(lèi)似,CPEBP 家族成員(如CPEBPβ) 亦能與IL4 啟動(dòng)子結(jié)合,是內(nèi)源性IL4 表達(dá)的潛在誘導(dǎo)因子,。同時(shí),CPEBP 亦能抑制IL22 和IFN-γRNA 的合成,但不影響其它Th2型細(xì)胞因子。JunB 屬于c-jun轉(zhuǎn)錄因子家族,選擇性表達(dá)于Th2 效應(yīng)細(xì)胞,能與AP21 特異結(jié)合,激活IL-4 轉(zhuǎn)錄,。JunB 介導(dǎo)的IL-4 表達(dá)需要JNK激酶和c2maf 的協(xié)作,并能誘導(dǎo)分化中的Th2 細(xì)胞產(chǎn)生IL-5,、IL-6 和IL-10。
4.1 JAK-STAT
JAK-STAT 是細(xì)胞因子受體介導(dǎo)的主要信號(hào)轉(zhuǎn)導(dǎo)通路,。IL-12 能通過(guò)STAT4 介導(dǎo)Th1 細(xì)胞分化, IL-4 可通過(guò)STAT6 介導(dǎo)Th2 細(xì)胞分化,。然而,STAT 介導(dǎo)的目的基因尚不清楚,在IL-4 啟動(dòng)子附近亦未發(fā)現(xiàn)STAT 結(jié)合位點(diǎn)。盡管STAT4 或STAT6 基因敲除實(shí)驗(yàn)顯示IFN-γ或IL-4依然正常分泌,但缺少STAT信號(hào)時(shí)GATA23,、T-bet 以及細(xì)胞因子的表達(dá)則大受影響,。推測(cè)STAT 與GATA23、c2maf ,、T2bet 等轉(zhuǎn)錄因子密切相關(guān),闡明這些內(nèi)在的有助于搞清Th 細(xì)胞分化的分子機(jī)制,。Bcl26 由于能與STAT6 競(jìng)爭(zhēng)相同的結(jié)合位點(diǎn),被認(rèn)為是Th2的負(fù)向調(diào)節(jié)因子。新近發(fā)現(xiàn)SOCS5 能通過(guò)與IL-4R 的相互作用,負(fù)向調(diào)節(jié)IL-4PSTAT6 通路,從而抑制Th2 分化,。
4.2 MAPK
哺乳動(dòng)物的MAPK信號(hào)通路包括細(xì)胞外信號(hào)調(diào)節(jié)激酶(ERK) ,、c-Jun N 端激酶(JNK)P應(yīng)激激活的蛋白激酶(SAPK) 、P38MAPK 以及ERK5PBMK1 四條途徑,。每條通路均通過(guò)保守的三級(jí)酶促級(jí)聯(lián)反應(yīng)[MAPKKK→MAPKK(MKK) →MAPK]激活轉(zhuǎn)錄因子,調(diào)節(jié)特定基因的表達(dá),。ERK位于癌基因Ras下游,Ras等不同激活信號(hào)通過(guò)Raf2MEK1 途徑激活ERK,通常稱(chēng)Ras2ERK通路,包括ERK1 和ERK2,。前已述及TCR 激活信號(hào)能激活MEKPERK,促使Th1分化。H2Ras 轉(zhuǎn)基因小鼠研究卻發(fā)現(xiàn),Ras2ERK通路經(jīng)非TCR 激活信號(hào)激活后,能上調(diào)JAK1激酶活性,促進(jìn)STAT6 酪氨酸磷酸化,并提示Ras-ERK與JAK-STAT通路之間存在交叉,??梢?jiàn),Ras2ERK通路與Th 細(xì)胞分化的確切關(guān)系尚待研究。
p38蛋白有4種亞型:p38-α,、p38-β,、p38-γ和p38-δ。MKK3,、MKK6 是其主要的蛋白激酶,應(yīng)激或炎癥刺激等情況下可被激活,。p38激活后的直接底物為MAPK激活的蛋白激酶(MAPKAPK)22P3 ,其目的基因啟動(dòng)子中含有CRE 或能與CREBPATF 和AP21 家族相互作用敏感的反應(yīng)元件。p382α,、MKK3P6等轉(zhuǎn)基因動(dòng)物實(shí)驗(yàn)發(fā)現(xiàn)抑制p38MAPK 通路的激活,IFN2γ水平明顯下降,。由于IFN2γ啟動(dòng)子近側(cè)和遠(yuǎn)側(cè)功能性活化元件處存在c-junPATF2 結(jié)合位點(diǎn)以及一系列其它ATF結(jié)合位點(diǎn),且近側(cè)IFN2γ元件攜帶c-junPATF2 結(jié)合位點(diǎn), 呈現(xiàn)Th1 特異性。因此,p38MAPK通路可能通過(guò)激活IFN2γ 轉(zhuǎn)錄,促使Th1細(xì)胞分化,。
JNK亦是一種應(yīng)激激活的蛋白激酶,包括JNK1P2P3,。激活的磷酸化級(jí)聯(lián)反應(yīng)為MEKK1 ,2 →MKK4PSEK1 →JNKPSAPK(MKK7 亦能激活JNK) 。JNKPSAPK能使c2jun,、ATF22 等磷酸化并增加其轉(zhuǎn)錄活性,促進(jìn)c-fos,、c-Jun、ATF22 調(diào)節(jié)基因的表達(dá),。JNK2 基因敲除研究發(fā)現(xiàn)小鼠IFN2γ水平降低,Th1 免疫應(yīng)答缺陷,但Th2 免疫應(yīng)答不受影響,。將IFN-γ 加入JNK2缺失的T 細(xì)胞中則能恢復(fù)上述缺陷,產(chǎn)生正常的Th1免疫應(yīng)答。JNK1 敲除的小鼠呈現(xiàn)強(qiáng)烈的Th2 免疫應(yīng)答,即使在Th1 條件下亦產(chǎn)生大量IL-4,、IL-5 和IL10 等Th2 型細(xì)胞因子,進(jìn)而發(fā)現(xiàn)JNK1 敲除小鼠核內(nèi)NF-AT2 水平上升,提示JNK1 能負(fù)向調(diào)節(jié)核內(nèi)NF2AT2 的水平,從而影響Th2 型細(xì)胞因子產(chǎn)生,。
4.3 NF-κBPI-κB
正常情況下,NF2κB 與其抑制因子I-κB 存在于胞漿中。隨著上游激酶(NF-κB誘導(dǎo)激酶或MEKK1) 的激活,I-κB 激酶活化并出現(xiàn)I-κB 蛋白磷酸化,導(dǎo)致它們與NF-κB 分離并降解,而游離的NF2κB則轉(zhuǎn)入胞核內(nèi)轉(zhuǎn)導(dǎo)激活目的基因,。NF-κB 因子在Th細(xì)胞分化中的作用研究目前剛起步,。已有研究發(fā)現(xiàn),抑制NF2κB 活性可阻斷GATA23 表達(dá)和Th2 型細(xì)胞因子產(chǎn)生。
5,、 結(jié)語(yǔ)
細(xì)胞分化過(guò)程本質(zhì)上是細(xì)胞基因的按序表達(dá)和選擇性表達(dá)的結(jié)果,因此,細(xì)胞分化調(diào)控的基本規(guī)律就是基因表達(dá)調(diào)節(jié),。對(duì)于Th 細(xì)胞分化而言,zui終表現(xiàn)為控制細(xì)胞應(yīng)答的有關(guān)基因的活化與表達(dá),細(xì)胞信號(hào)轉(zhuǎn)導(dǎo)通路在此起了重要的中介作用。近幾年,Th 細(xì)胞分化機(jī)制研究進(jìn)展迅速,已認(rèn)識(shí)到TCR 活化信號(hào),、輔助受體以及細(xì)胞因子環(huán)境等因素共同介導(dǎo)了Th1PTh2 細(xì)胞的分化方向,并逐步深入到轉(zhuǎn)錄因子,、信號(hào)轉(zhuǎn)導(dǎo)、基因調(diào)控水平,。我們相信,Th 細(xì)胞分化的復(fù)雜性,、調(diào)節(jié)的性以及信號(hào)通路和相關(guān)因子的交叉性等問(wèn)題的zui終闡明,必將給免疫性疾病的藥物治療帶來(lái)重大突破。

相關(guān)通路指標(biāo):
T-Cell Activation:
Regulators of T-Cell Activation: CD2, CD276, CD47, DPP4, CD3D, CD3E, CD3G, CD4, CD7, CD80, CD86, CD8A, CD8B, FOXP3, ICOSLG, IRF4, LAG3, LCK, MAP3K7 (TAK1), MICB, NCK1, TNFSF14, VAV1.
T-Cell Proliferation: CD28, CD3E, ICOSLG, IL1B, IL10, IL12B, IL18, NCK1, RIPK2, TNFSF14.
T-Cell Differentiation: ADA, APC, BCL2, BLM, CD1D, CD2, CD27 (TNFRSF7), CD4, CD80, CD86, EGR1, IL12B, IL15, IL2, IRF4, NOS2 (iNOS), PTPRC, SOCS1.
T-Cell Polarization: CCL3, CCR1, CCR2, CCR3, CCR4, CCR5, CD274, CD28, CD4, CD40LG (TNFSF5), CSF2 (GM-CSF), CXCR3, CXCR4, IFNG, IL12A, IL12RB1, IL12RB2, IL18R1, IL2, IL4, IL4R, IL5, TGFB1.
Regulators of Th1 and Th2 Development: CD2, CD40 (TNFRSF5), CD5, CD7, CSF2 (GM-CSF), IFNG, IL10, IL12A, IL13, IL3, IL4, IL5, TLR2, TLR4, TLR9.
Th1 & Th2 Differentiation: CD28, CD40 (TNFRSF5), CD40LG (TNFSF5), CD86, IFNG, IL12A, IL12B, IL12RB1, IL12RB2, IL18, IL18R1, IL2, IL2RA, IL4, IL4R, IL6.
B-Cell Activation:
Antigen Dependent B-cell Activation: CD28, CD4, CD40 (TNFRSF5), CD40LG (TNFSF5), CD80, FAS (TNFRSF6), FASLG (TNFSF6), IL10, IL2, IL4.
Other Genes involved in B-Cell Activation: ADA, CXCR5, ICOSLG, IL6, IL7, MS4A1, TGFB1.
B-Cell Proliferation: BCL2, CD27 (TNFRSF7), CD40 (TNFRSF5), CD81, IL10, IL7, PTPRC.
B-Cell Differentiation: ADA, AICDA, BLNK, CD27 (TNFRSF7), IL10, IL11, IL4, RAG1.
Other Immune Cell Activation:
Macrophage Activation: IL13, IL4, TLR1, TLR4, TLR6.
Neutrophil Activation: IL8.
Natural Killer Cell Activation: CD2, IL12A, IL12B, IL2.
Leukocyte Activation: CX3CL1.
通路研究工具:
?細(xì)胞譜系鑒定系列 | PCR Array of Cell Lineage Identification | HU-CLI-084 |
胰島素抵抗系列 | PCR Array of Insulin Resistance | HU-INR-084 |
T細(xì)胞和B細(xì)胞激活系列 | PCR Array of T-Cell & B-Cell Activation | HU-TCA-084 |
終末分化指標(biāo)系列 | PCR Array of Terminal Differentiation Markers | HU-TDM-084 |
PCR Array:

Q-PCR array是信號(hào)通路基礎(chǔ)生物學(xué)研究和臨床疾病研究的重要工具,。這種功能分類(lèi)芯片運(yùn)用成熟的SYBR Green熒光定量PCR技術(shù),,專(zhuān)注于一個(gè)信號(hào)通路中基因的表達(dá)水平檢測(cè)。與傳統(tǒng)的高密度表達(dá)譜芯片相比,,Q-PCR array具有針對(duì)性強(qiáng),、靈敏度高、準(zhǔn)確可靠等優(yōu)點(diǎn),。 芯片上的基因包括了與研究對(duì)象有確定關(guān)系的基因,,或至少是與研究對(duì)象的關(guān)系有待考證的基因。如果研究對(duì)象是某一生物學(xué)通路的基因,,則使用針對(duì)該類(lèi)基因的功能分類(lèi)基因芯片比使用高密度表搭配芯片更加有效便利,。
BioTNT QPCR芯片特點(diǎn):
l 靈敏度高,樣本使用量低,;
l 線(xiàn)性范圍廣,,可同時(shí)檢測(cè)表達(dá)水平差異大的基因;
l 每個(gè)基因的檢測(cè)引物優(yōu)化后保證高擴(kuò)增率和產(chǎn)物單一,;
l 重復(fù)性高,,Ct值的平均差異只有0.25個(gè)循環(huán);
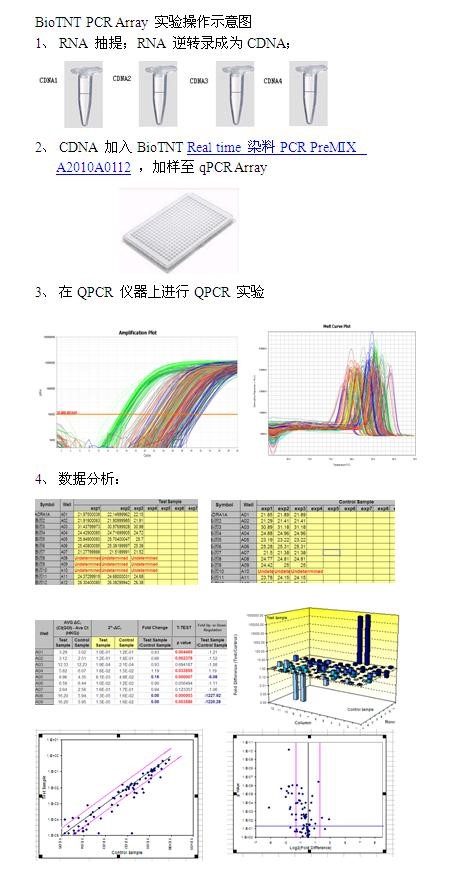
BioTNT QPCR Array芯片操作流程:
相關(guān)產(chǎn)品




免責(zé)聲明
- 凡本網(wǎng)注明“來(lái)源:化工儀器網(wǎng)”的所有作品,,均為浙江興旺寶明通網(wǎng)絡(luò)有限公司-化工儀器網(wǎng)合法擁有版權(quán)或有權(quán)使用的作品,,未經(jīng)本網(wǎng)授權(quán)不得轉(zhuǎn)載、摘編或利用其它方式使用上述作品,。已經(jīng)本網(wǎng)授權(quán)使用作品的,,應(yīng)在授權(quán)范圍內(nèi)使用,并注明“來(lái)源:化工儀器網(wǎng)”,。違反上述聲明者,,本網(wǎng)將追究其相關(guān)法律責(zé)任。
- 本網(wǎng)轉(zhuǎn)載并注明自其他來(lái)源(非化工儀器網(wǎng))的作品,,目的在于傳遞更多信息,,并不代表本網(wǎng)贊同其觀點(diǎn)和對(duì)其真實(shí)性負(fù)責(zé),不承擔(dān)此類(lèi)作品侵權(quán)行為的直接責(zé)任及連帶責(zé)任,。其他媒體,、網(wǎng)站或個(gè)人從本網(wǎng)轉(zhuǎn)載時(shí),必須保留本網(wǎng)注明的作品第一來(lái)源,,并自負(fù)版權(quán)等法律責(zé)任,。
- 如涉及作品內(nèi)容、版權(quán)等問(wèn)題,,請(qǐng)?jiān)谧髌钒l(fā)表之日起一周內(nèi)與本網(wǎng)聯(lián)系,,否則視為放棄相關(guān)權(quán)利。


 采購(gòu)中心
采購(gòu)中心