 化工儀器網(wǎng)
化工儀器網(wǎng)
技術文章
研究闡述組蛋白修飾H2AK119ub1在染色質上蔓延以及跨細胞周
閱讀:787 發(fā)布時間:2020-3-30生物大分子國家重點實驗室李國紅課題組研究系統(tǒng)闡述了組蛋白修飾H2AK119ub1在染色質上蔓延以及跨細胞周期繼承的具體機制。
真核細胞內,,染色質是表觀遺傳信息的主要調控界面,。諸多表觀遺傳因子通過調節(jié)染色質結構的開放與關閉,控制了基因的表達與沉默,,從而維持了不同譜系細胞內基因的差異表達,。其中,多梳抑制復合物1(PRC1)是一類重要的表觀遺傳調控因子,,它主要通過壓縮染色質結構和催化形成H2AK119ub1修飾這兩種途徑促進了兼性異染色質的形成,,并以此維持了PRC1靶基因的沉默狀態(tài)。H2AK119ub1在染色質局部往往連綿不斷形成非常寬廣的H2AK119ub1區(qū)域,,但是其產生的機制一直未被解析,。
課題組結合核小體pull-down、體外泛素化,、TetR-TetO靶向和CHIP-seq等實驗發(fā)現(xiàn)RYBP和YAF2蛋白能通過ZNF結構域特異識別并結合H2AK119ub1,,借此使RYBP-PRC1和YAF2-PRC1復合物與其催化產物之間形成所謂的“酶-產物之間的正反饋機制”,從而促進了H2AK119ub1向周邊的蔓延,。
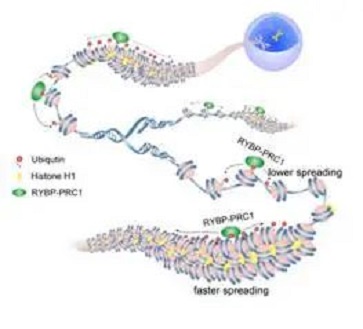
有趣的是,,通過基于組蛋白修飾和DNaseI高敏位點的染色質狀態(tài)分析,發(fā)現(xiàn)H2AK119ub1主要分布在結構緊密的異染色質區(qū)域,,因此提出非常規(guī)PRC1介導的H2AK119ub1蔓延也可能受到染色質高級結構的調控,。之前,李國紅和朱平課題組合作,,利用冷凍電鏡技術解析了組蛋白H1壓縮30nm染色質纖維的高精度結構,,揭示染色質纖維是以四核小體為結構單元的左手雙螺旋結構,并發(fā)現(xiàn)相間核小體之間的相互作用在維系四核小體內部和四核小體之間的穩(wěn)定性中發(fā)揮了重要作用,。
因此,,30nm染色質纖維可能通過核小體-核小體配對(nucleosome-pairing)的方式促進了非常規(guī)PRC1介導的H2AK119ub1在染色質纖維上的蔓延?;谠摷僭O,,作者進行了進一步的實驗并發(fā)現(xiàn)H1壓縮的30nm染色質纖維的確促進了H2AK119ub1向鄰近染色質區(qū)域的蔓延。
在體外泛素化實驗中,,通過引入組蛋白相應位置的突變體,,證明H1對H2AK119ub1蔓延的促進作用主要通過改變染色質高級結構來實現(xiàn)的。并且,,根據(jù)30nm染色質纖維的電鏡結構,,作者推測這種促進機制可能遵循相間核小體-核小體的配對原則,。
*,DNA復制過程中染色質會經歷一次“破壞-重建”的過程,。在DNA子鏈上新合成組蛋白的裝配會導致組蛋白修飾至少稀釋一倍,,如果沒有有效的組蛋白修飾恢復機制,細胞身份很難得以維持,。
利用TetR-TetO靶向系統(tǒng),,作者發(fā)現(xiàn)單純靶向RING1B在8xTetO附近僅能形成很窄的H2AK119ub1修飾,當利用多西環(huán)素(Doxcycline,,Dox)洗去RING1B-TetR-HA時,,異位形成的H2AK119ub1修飾并不能在子代細胞內得到有效的維持。然而過表達RYBP促進H2AK119ub1向兩側蔓延形成大約10kb的區(qū)域時,,實驗發(fā)現(xiàn)正反饋機制能介導H2AK119ub1在8xTetO附近維持至少6個細胞周期,。
同時發(fā)現(xiàn)H1介導的染色質壓縮對于H2AK119ub1的遺傳也至關重要。另外在全基因組上,,發(fā)現(xiàn)RYBP突變和H1c/H1d/H1e三敲除導致的H2AK119ub1降低以及基因表達變化有很大程度的重疊,。并且利用神經分化模型,發(fā)現(xiàn)RYBP突變和H1c/H1d/H1e三敲除均導致神經前體細胞分化障礙,。
因此,,說明RYBP-/YAF2-PRC1復合物和組蛋白H1通過協(xié)同促進細胞周期間H2AK119ub1的播散參與了細胞分化過程,提示該表觀遺傳通路具有重要的生物學功能,。