

 QQ交談
QQ交談 MSN交談
MSN交談請輸入產(chǎn)品關(guān)鍵字:

郵編:201100
聯(lián)系人:陳
電話:021-64133189
傳真:021-64129208
手機(jī):13788995069
留言:發(fā)送留言
個性化:www.runwelltac.com
網(wǎng)址:www.runwelltac.com
商鋪:http://sorrent.com.cn/st111352/
半乳糖凝集素3在血管生成中的作用
點(diǎn)擊次數(shù):3596 發(fā)布時間:2011-11-7
Galectin-3在血管生成中的作用
半乳糖凝集素Galectin-3在血管生成中的作用
www.biomart.cn/runwell/index.htm
某些生理過程,,如胚胎發(fā)育,、生殖功能和傷口愈合,都需要新血管的形成,。從現(xiàn)有的血管形成新血管的細(xì)胞機(jī)制被稱為血管生成,。血管生成是一個多步驟的過程,包括一些細(xì)胞因子(peprotech)和生長因子的協(xié)調(diào)行動,,對血管內(nèi)皮細(xì)胞發(fā)揮趨化,、促有絲分裂和調(diào)節(jié)作用1。
研究表明,,碳水化合物結(jié)合蛋白以及它們各自的糖綴合物配體在血管生成中也扮演了重要角色,。2,3 具體來說,近期的研究表明Galectin-3 促進(jìn)了血管生成,。4,5 Galectin-3 是半乳糖凝集素(Galectin)家族中的一員,,能與包含N- 乙酰乳糖胺的糖蛋白特異結(jié)合。6 所有的半乳糖凝集素在C 端至少含有一個糖識別域(CRD),。然而,,Galectin-3 卻是家族中*一個成員,包含了一個與富含脯氨酸,、甘氨酸和酪氨酸的N 端結(jié)構(gòu)域連接的CRD,。Galectin-3 的N 端部分介導(dǎo)了配體結(jié)合時的寡聚化。6 Galectin-3 主要分布在胞質(zhì)中,,但也能分泌到胞外發(fā)揮作用,,在胞外它與包含β - 半乳糖苷的細(xì)胞表面糖綴合物結(jié)合。
在研究表明Galectin-3 影響了人臍靜脈內(nèi)皮細(xì)胞(HUVEC)的趨化和分化之后,,它在血管生成中的作用才被發(fā)現(xiàn),。在體外,Galectin-3 是趨化的,,并誘導(dǎo)了HUVEC 中毛細(xì)血管的形成,。4 Galectin-3 還促進(jìn)了體內(nèi)的血管生成。4,5 這些過程依賴Galectin-3 的糖類識別和寡聚化,。
zui近,,Markowska 等的一篇文章闡明了Galectin-3 介導(dǎo)血管生成的可能機(jī)制。5 這些作者發(fā)現(xiàn),Galectin-3 參與了血管內(nèi)皮生長因子(VEGF)和堿性成纖維細(xì)胞生長因子(bFGF)誘導(dǎo)的血管生成,。通過小干擾RNA 降低HUVEC 中Galectin-3 的表達(dá),,減少了VEGF 和bFGF 介導(dǎo)的遷移和毛細(xì)血管形成。5 此外,,在Galectin-3 缺陷型小鼠中,,兩種生長因子都未能誘導(dǎo)新生血管形成。5 將HUVEC 裂解液蛋白用Galectin-3 親和柱分離后進(jìn)行質(zhì)譜分析,,發(fā)現(xiàn)整合素αVβ3 作為結(jié)合伙伴,,可能參與了Galectin-3 介導(dǎo)的血管生成。5 一般來說,,整合素成簇引發(fā)了細(xì)胞間信號分子的激活,,如粘附斑激酶(FAK)。7 具體來說,,研究表明整合素αVβ3 參與了生長因子介導(dǎo)的血管生成,。7-10 Markowska 及其同事證明,用Galectin-3 處理HUVEC,,促進(jìn)了整合素αVβ3 的成簇和FAK 的激活,。5 此外,用整合素αVβ3 的抗體預(yù)處理則抑制了Galectin-3 誘導(dǎo)的HUVEC 遷移和毛細(xì)血管形成,。
他們假設(shè),,Galectin-3 與整合素αVβ3 上存在的復(fù)雜N 聚糖相互作用,激活了影響VEGF 和bFGF 介導(dǎo)血管生成的信號通路,。作者生成了表達(dá)整合素αVβ3 的HUVEC,,而高親和力的Galectin-3配體(β1,6-N- 乙酰氨基葡萄糖(GlcNAc)分支的N 聚糖)的表達(dá)下降。他們用攜帶有針對人N 乙酰氨基葡萄糖轉(zhuǎn)移酶V(GnTV)的小發(fā)夾RNA(shRNA)的慢病毒轉(zhuǎn)染HUVEC,,實(shí)現(xiàn)了這一點(diǎn),,因GnTV 合成了N 聚糖上的β1-6GlcNAc 分支。5,11,12 干擾HUVEC 中GnTV 的表達(dá)減弱了Galectin-3 與細(xì)胞表面糖蛋白的結(jié)合,。5 這些轉(zhuǎn)染了GnTV shRNA 的細(xì)胞在暴露于Galectin-3,、VEGF 或bFGF時未遷移或形成血管結(jié)構(gòu)。5 此外,,Galectin-3 和兩種生長因子都未能促進(jìn)活體GnTV–/– 小鼠內(nèi)的血管生成,。
除了需要血管生成的關(guān)鍵生理過程,許多疾病狀態(tài)也引發(fā)了異常的毛細(xì)管生長,。4 例如,,新血管形成對癌細(xì)胞的生長、侵襲和轉(zhuǎn)移很關(guān)鍵,。1 研究發(fā)現(xiàn),,癌癥患者中的Galectin-3 血清水平比正常對照要高,。13 Galectin-3 調(diào)節(jié)血管生成的細(xì)胞機(jī)制的闡明提供了一條新的途徑,有可能從藥理上調(diào)控病理性新血管生成,。
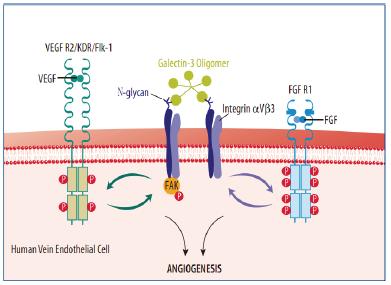
在生長因子介導(dǎo)的血管生成中,,Galectin-3 激活整合素αVβ3。Galectin-3,,一種β 半乳糖苷結(jié)合蛋白,,曾被認(rèn)為參與了血管生成;然而,,Galectin-3 所采用的機(jī)制仍不清楚,。Markowska 及其同事證明,Galectin-3 與整合素αVβ3 上的復(fù)雜N 聚糖結(jié)合,,引發(fā)了整合素成簇和FAK 激活,。5 激活的整合素αVβ3 隨后影響了VEGF 和bFGF 誘導(dǎo)的血管生成,。此項(xiàng)研究*大量研究空缺,,得出血管生成活性依賴整合素與生長因子受體的協(xié)調(diào)作用。在配體結(jié)合時,,整合素αVβ3,,特別是β3 亞基,與VEGF R2 和FGF R1 形成復(fù)合物,。9,10,14 這種整合素αVβ3 與VEGF R2 之間的相互作用對VEGF 誘導(dǎo)的VEGF R2 磷酸化以及VEGF R2 信號通路的激活很關(guān)鍵,。7,9,10 在bFGF 誘導(dǎo)的血管生成中,整合素αVβ3 也是ERK1/2 持續(xù)激活所必需的,。15 研究表明,,生長因子以相似的方式調(diào)節(jié)了整合素發(fā)揮作用。VEGF 和FGF 受體激活增加了整合素αVβ3 對其配體的親和力,。