





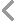


手持式葉綠素?zé)晒鈨x---FluorPen FP110
- 公司名稱 因子科學(xué)儀器(山東)有限公司
- 品牌 其他品牌
- 型號(hào)
- 產(chǎn)地
- 廠商性質(zhì)
- 更新時(shí)間 2021/3/5 16:14:04
- 訪問次數(shù) 432
聯(lián)系我們時(shí)請(qǐng)說明是化工儀器網(wǎng)上看到的信息,,謝謝!
| 產(chǎn)地類別 | 國產(chǎn) | 價(jià)格區(qū)間 | 面議 |
|---|---|---|---|
| 應(yīng)用領(lǐng)域 | 醫(yī)療衛(wèi)生,生物產(chǎn)業(yè) |
手持式葉綠素?zé)晒鈨x---FluorPen FP110用于實(shí)驗(yàn)室、溫室和野外快速測量植物葉綠素?zé)晒鈪?shù),,具有便攜性強(qiáng),、精度高、性價(jià)比高等特點(diǎn),;雙鍵操作,,具圖形顯示屏,內(nèi)置鋰電和數(shù)據(jù)存儲(chǔ),廣泛應(yīng)用于研究植物的光合作用,、脅迫監(jiān)測,、除草劑檢測或突變體篩選,還可用于生物毒理的生物檢測,,如通過不同植物對(duì)土壤或水質(zhì)污染的葉綠素?zé)晒忭憫?yīng),,找出敏感植物作為生物傳感器用于生物檢測。FP110配備多種葉夾型號(hào),,用于不同的樣品與研究,。
應(yīng)用領(lǐng)域
手持式葉綠素?zé)晒鈨x---FluorPen FP110適用于光合作用研究和教學(xué),植物及分子生物學(xué)研究,,農(nóng)業(yè),、林業(yè),生物技術(shù)領(lǐng)域等,。研究內(nèi)容涉及光合活性,、脅迫響應(yīng)、農(nóng)藥藥效測試,、突變篩選等,。
- 植物光合特性研究
- 光合突變體篩選與表型研究
- 生物和非生物脅迫的檢測
- 植物抗脅迫能力或者易感性研究
- 農(nóng)業(yè)和林業(yè)育種、病害檢測,、長勢與產(chǎn)量評(píng)估
- 除草劑檢測
- 教學(xué)
功能特點(diǎn)
- 結(jié)構(gòu)緊湊,、便攜性強(qiáng),LED光源,、檢測器,、控制單元集成于僅手機(jī)大小的儀器內(nèi),重量僅188g
- 功能強(qiáng)大,,是葉綠素?zé)晒饧夹g(shù)的高新結(jié)晶產(chǎn)品,,具備了大型熒光儀的所有功能,可以測量所有葉綠素?zé)晒鈪?shù)
- 內(nèi)置了所有通用葉綠素?zé)晒夥治鰧?shí)驗(yàn)程序,,包括3套熒光淬滅分析程序,、3套光響應(yīng)曲線程序、OJIP快速熒光動(dòng)力學(xué)曲線等
- 高時(shí)間分辨率,,可達(dá)10萬次每秒,,自動(dòng)繪出OJIP曲線并給出26個(gè)OJIP–test參數(shù)
- FluorPen專業(yè)軟件功能強(qiáng)大,可下載,、展示葉綠素?zé)晒鈪?shù)圖表,,也可以通過軟件直接控制儀器進(jìn)行測量
- 具備無人值守自動(dòng)監(jiān)測功能
- 內(nèi)置藍(lán)牙與USB雙通訊模塊,GPS模塊,,輸出帶時(shí)間戳和地理位置的葉綠素?zé)晒鈪?shù)圖表
- 配備多種葉夾型號(hào):固定葉夾式(適于實(shí)驗(yàn)室內(nèi)暗適應(yīng)或夜間快速測量)、分離葉夾式(適用于野外暗適應(yīng)測量)、探頭式(透明光纖探頭,,具備葉片固定裝置,,用于非接觸性測量監(jiān)測或光適應(yīng)條件下的葉綠素?zé)晒獗O(jiān)測)、用戶定制式等
- 可選配野外自動(dòng)監(jiān)測式熒光儀,,防水防塵設(shè)計(jì)
測量程序與功能
- Ft:瞬時(shí)葉綠素?zé)晒?暗適應(yīng)完成后Ft=F0
- QY:量子產(chǎn)額,,表示光系統(tǒng)II 的效率,等于Fv/Fm(暗適應(yīng)狀態(tài))或ΦPSII (光適應(yīng)狀態(tài)),。
- OJIP:快速熒光動(dòng)力學(xué)曲線,,用于研究植物暗適應(yīng)后的快速熒光動(dòng)態(tài)變化
- NPQ:熒光淬滅動(dòng)力學(xué)曲線,用于研究植物從暗適應(yīng)到光適應(yīng)狀態(tài)的熒光淬滅變化過程,。
- LC:光響應(yīng)曲線,,用于研究植物對(duì)不同光強(qiáng)的熒光淬滅反應(yīng)。
- PAR:光合有效輻射,,測量環(huán)境中植物生長可以利用的400-700nm實(shí)際光強(qiáng)(限PAR型號(hào)),。
應(yīng)用案例
2017年4月,美國國家航空*(NASA)新一代*植物培養(yǎng)器(Advanced Plant Habitat,,APH)搭載聯(lián)盟號(hào)MS-04貨運(yùn)飛船抵達(dá)空間站,。宇航員使用FluorPen手持儀葉綠素?zé)晒鈨x在其中開展植物生理學(xué)及太空食物種植(growth of fresh food in space)的研究。
參考文獻(xiàn)
- F Dang, et al. 2019. Discerning the Sources of Silver Nanoparticle in a Terrestrial Food Chain by Stable Isotope Tracer Technique. Environmental Science & Technology 53(7): 3802-3810
- N Moghimi, et al. 2019. New candidate loci and marker genes on chromosome 7 for improved chilling tolerance in sorghum. Journal of Experimental Botany 70(12): 3357–3371
- M Rafique, et al. 2019. Potential impact of biochar types and microbial inoculants on growth of onion plant in differently textured and phosphorus limited soils. Journal of Environmental Management 247: 672-680
- P Soudek, et al. 2019. Thorium as an environment stressor for growth of Nicotiana glutinosa plants. Environmental and Experimental Botany 164: 84-100
- JA Pérez-Romero, et al. 2019. Investigating the physiological mechanisms underlying Salicornia ramosissima response to atmospheric CO2 enrichment under coexistence of prolonged soil flooding and saline excess. Plant Physiology and Biochemistry 135: 149-159
- D Shao, et al. 2019. Physiological and biochemical responses of the salt-marsh plant Spartina alterniflora to long-term wave exposure. Annals of Botany, DOI: 10.1093/aob/mcz067
- C Cirillo, et al. 2019. Biochemical, Physiological and Anatomical Mechanisms of Adaptation of Callistemon citrinus andViburnum lucidum to NaCl and CaCl2 Salinization. Front. Plant Sci. 10:742
- T Savchenko, et al. 2019. Waterlogging tolerance rendered by oxylipin-mediated metabolic reprogramming in Arabidopsis. Journal of Experimental Botany 70(10): 2919–2932
- M Liu, et al. 2019. Strong turbulence benefits toxic and colonial cyanobacteria in water: A potential way of climate change impact on the expansion of Harmful Algal Blooms. Science of The Total Environment 670: 613-622
- PK Tiwari, et al. 2019. Liquid assisted pulsed laser ablation synthesized copper oxide nanoparticles (CuO-NPs) and their differential impact on rice seedlings. Ecotoxicology and Environmental Safety 176: 321-329
- JA Pérez-Romero, et al. 2018. Atmospheric CO2 enrichment effect on the Cu-tolerance of the C4 cordgrass Spartina densiflora. Journal of Plant Physiology 220: 155-166
- SK Yadav, et al. 2018. Physiological and Biochemical Basis of Extended and Sudden Heat Stress Tolerance in Maize. Proceedings of the National Academy of Sciences 88(1): 249-263
- D Balfagón, et al. 2018. Involvement of ascorbate peroxidase and heat shock proteins on citrus tolerance to combined conditions of drought and high temperatures. Plant Physiology and Biochemistry 127: 194-199
- JI Vílchez, et al. 2018. Protection of Pepper Plants from Drought by Microbacterium sp. 3J1 by Modulation of the Plant's Glutamine and α-ketoglutarate Content: A Comparative Metabolomics Approach. Front. Microbiol. 9: 284
- MC Sorrentino, et al. 2018. Performance of three cardoon c*rs in an industrial heavy metal-contaminated soil: Effects on morphology, cytology and photosynthesis. Journal of Hazardous Materials 351: 131-137
- E Niewiadomska, et al. 2018. Lack of tocopherols influences the PSII antenna and the functioning of photosystems under low light. Journal of Plant Physiology 223: 57-64
- S Singh, et al. 2018. Cadmium toxicity and its amelioration by kinetin in tomato seedlings vis-à-vis ascorbate-glutathione cycle. Journal of Photochemistry and Photobiology B: Biology 178: 76-84
- EL Fry, et al. 2018. Drought neutralises plant–soil feedback of two mesic grassland forbs. Oecologia 186(4): 1113–-125
附:OJIP參數(shù)及計(jì)算公式
Bckg = background
Fo: = F50µs; fluorescence intensity at 50 µs
Fj: = fluorescence intensity at j-step (at 2 ms)
Fi: = fluorescence intensity at i-step (at 60 ms)
Fm: = maximal fluorescence intensity
Fv: = Fm - Fo (maximal variable fluorescence)
Vj = (Fj - Fo) / (Fm - Fo)
Fm / Fo = Fm / Fo
Fv / Fo = Fv / Fo
Fv / Fm = Fv / Fm
Mo = TRo / RC - ETo / RC
Area = area between fluorescence curve and Fm
Sm = area / Fm - Fo (multiple turn-over)
Ss = the smallest Sm turn-over (single turn-over)
N = Sm . Mo . (I / Vj) turn-over number QA
Phi_Po = (I - Fo) / Fm (or Fv / Fm)
Phi_o = I - Vj
Phi_Eo = (I - Fo / Fm) . Phi_o
Phi_Do = 1 - Phi_Po - (Fo / Fm)
Phi_Pav = Phi_Po - (Sm / tFM); tFM = time to reach Fm (in ms)
ABS / RC = Mo . (I / Vj) . (I / Phi_Po)
TRo / RC = Mo . (I / Vj)
ETo / RC = Mo . (I / Vj) . Phi_o)
DIo / RC = (ABS / RC) - (TRo / RC)


 采購中心
采購中心
 化工儀器網(wǎng)
化工儀器網(wǎng)